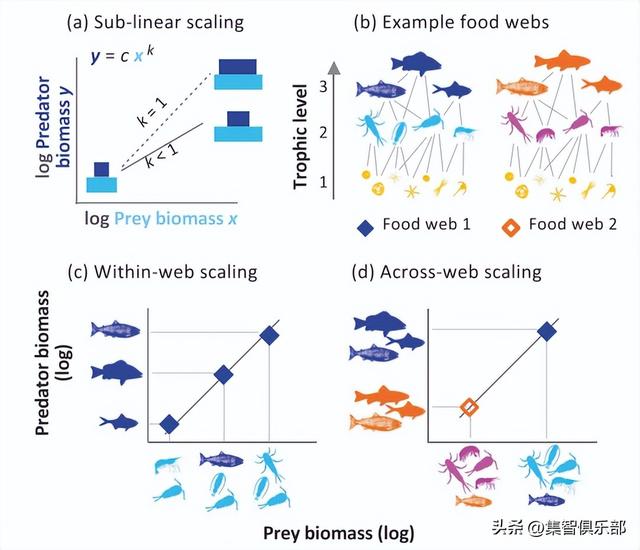
捕食者与被捕食的生物量比例是营养结构的关键因素,通常科学家们都是从食物链的视角对此展开研究,而不关注能量传递的渠道(比如是否为杂食),其实能量传递渠道可能会对群落结构起支配作用。近日发表于 Nature Communications 的一项新研究,通过描述141个淡水、海洋和陆地食物网的生物量结构补足了这一缺陷,这些食物网跨越了广阔的群落生物量阶梯(gradient in community biomass)。
本文开发了在复杂食物网中计算捕食者和被捕食者生物量的方法,检验了捕食者和被捕食者的生物量的亚线性规模比例关系。结果确实发现了在复杂的食物网中依然存在一个统一的亚线性规模法则:在食物网内(within food webs),捕食者的总生物量与它们的猎物总生物量的比例接近3/4的幂指数。并且在食物网之间(across food webs),捕食者的总生物量和食物网中所有猎物的总生物量之间也出现了类似的亚线性规模法则,幂指数在不同类型生态系统中都表现出了令人惊讶的一致性。
研究领域:规模法则,食物网,生态系统

原文标题:
Consistent predator-prey biomass scaling in complex food webs
原文链接:
https://www.nature.com/articles/s41467-022-32578-5
1. 捕食者-猎物生物量的3/4规模法则理解驱动生态系统的结构和功能是生态学的一个基本目标。捕食者与猎物的生物量比例是衡量营养结构和群落动态的一个关键指标,并且与许多生态系统功能和服务相关联。然而,由于缺乏经验数据,负责驱动这些功能和规律的的主要机制仍然是不确定的。
之前发现指出了一个令人惊奇的规律:在水生和陆地系统中,捕食者与被捕食者生物量的比例,在生物量阶梯上都会发生变化。食肉动物生物量(例如,狮子、鬣狗和其他大型食肉动物的总生物量)在双对数尺度上与它们猎物的生物量(如水牛和其他食草动物)以亚线性的方式成比例关系,并且幂指数接近3/4。这意味着随着猎物生物量的增加,生物量金字塔会变得更有系统地偏重于底部(如图1a)。
有两个原因可以解释这种亚线性规模法则:(1)生物质量和生物生长的异速约束关系,这可能会导致群落的生物量亚线性缩放。(2)密度依赖效应:生物的质量在生物量阶梯上变化不大,亚线性缩放的原因在于不同生物阶梯的密度不同。
图1. 不同层次的捕食者-被捕食者生物量规模法则示意图。
a. 捕食者-被捕食者的幂指数k描述了金字塔形猎物生物量阶梯的相对变化。k=1说明没有相对变化。b. 自然界中的营养相互作用产生了一个复杂的相互作用网络,消费者的营养高度可以是一个非整数值。c. 在食物网内的规模法则。每个数据点代表在一个单一的食物网内,不同捕食者类群的生物量与它们的猎物的总生物量。不同的点代表同一张食物网中不同的捕食者。d. 食物网间的规模法则。每个数据点表示一个食物网内所有捕食者的总生物量,与所有被捕食者的总生物量。不同的点代表不同的食物网。在所有的案例中,使用各物种的丰度和平均体重来计算营养生物量。
然而,自然群落并没有被营养结构(如食草动物,食肉动物)形成的食物网很清晰地概念化。大多数的猎物是由许多捕食者共享的(如图1b),这会形成一张复杂的食物网,能量可以通过多种渠道传递。因此,与食物网复杂性相关的各种过程可能会改变捕食者-被捕食者的幂律关系。那么,一般的捕食者-被捕食者幂律是否适用于复杂的食物网呢?
本文开发了一个在复杂的食物网中定量刻画捕食者-被捕食者生物量的框架,计算了网络中每个分类单元的生物量,以及每个捕食者所有猎物的总生物量。同时也测试了杂食性程度和捕食者-被捕食者质量比(predator-prey body mass ratios,PPmR)的变化如何影响生物量的规模法则现象。本文使用的数据横跨不同的生态系统(淡水,海水,陆地三大生态系统),以及不同的营养层级,提供了一个机会来验证捕食-被捕食幂律关系的普遍性。
具体来说,本文试图回答这么几个问题:
(1) 捕食-被捕食关系生物量的规模法则在复杂食物网内部、食物网之间,分别是什么样的?是否在不同层级的生物组织中存在普适的幂律关系?
(2)杂食性生物和捕食-被捕食生物量比例在规模法则中扮演什么样的角色?
(3)捕食者和猎物生物量之间的联系是由猎物密度决定的,还是猎物平均质量的变化决定的?
2. 食物网内的生物量规模法则如图1c所示,本文为每个食物网构建了捕食与被捕食的生物量,使用线性混合效应模型(Linear mixed-effects model, LME)来决定标度指数k。发现无论是什么类型的生态系统,食物网的生物量的平均标度指数k都小于1,即是亚线性的,但具体的值在不同的生态系统中的置信区间范围差异较大(如图2)。
图2. 对于不同的生态系统类型,食物网内的捕食者-被捕食者生物量的规模法则。
a-c中每个数据点是在一个食物网内,不同的捕食者生物量和其猎物的总生物量之间的关系(虚线代表1:1的关系)。拟合的线代表平均的幂律指数。图中的文字代表对应系统的规模和信息。d-f.食物网的幂指数分布。实线是每个系统的平均值,在统计上三个系统都接近3/4。虚线表示95%置信区间。
值得注意的是,将拟合模型从最复杂的情况开始考虑,即假设每个生态系统类型都有不同的指数,一直到零模型(null model)
,即假设所有的系统都有相同的指数,对比之后发现,证明不同系统类型的指数具有显著差异的证据其实是非常弱。所以,在一个生态系统内,捕食者-被捕食者生物量的标度可以用平均指数描述
= 0.71
。这意味着,被捕食者生物量翻倍,仅带来捕食者生物量约3/4倍的增加。
另外,为了调查捕食者-被捕食者生物量比例(PPmR)和生物杂食性在这一规模关系中扮演的角色,本文还进行了进一步的分析,将这些变量作为额外固定效应项加入混合效应模型。这里将生物的杂食性定义为其捕食的物种在不同营养层级的方差大小。这些固定效应的加入增加了模型的解释性,并且这分析也解释了对于每个生态类型来说,捕食者生物量残差都会显著随着PPmR增加(如图3a-c)。
图3. 对于三个生态系统,变量对于捕食者生物量残差方差的解释性。
数据点是是使用线性混合效应模型对于食物网内的规模法则分析之后的捕食者生物量的残差。a-c是残差和捕食者-被捕食者生物量比例(PPmR)的关系。d-f是残差和捕食者杂食性的关系。捕食者生物量和PPmR之间有明显的正向关系(a-c),并且也和杂食性在土壤生态系统中成正向关系(f),但和杂食性并没有在淡水或者海洋环境中表现出相关性(d,e)。
3. 食物网间的生物量规模法则文章还探索了捕食者-被捕食者生物量的规模法则是否存在于食物网间。将每一个食物网内的所有捕食者加总,所有被捕食者加总,然后观察所有食物网的关系。这个跨食物网的分析在不同生态系统中揭示了令人惊讶的一致性——在淡水、海洋和陆地系统中标度指数分别为0.66,0.65,0.67,我们也能得到平均指数0.66,和在食物网内观察到的规模法则也很类似(即上文所说的0.71)。
进一步,文章还调查了猎物平均体重和猎物密度这两个变量在这一规模法则中起到的作用如何,发现猎物的平均质量和猎物总生物量无关。所以本文得出的结论是,这里观察到的捕食者生物量与猎物生物量的一致变化主要与猎物密度的变化有关,与猎物的平均大小无关。
4. 亚线性规模法则为何普遍存在?本文提供了一个在复杂食物网中对于捕食者-被捕食者生物量规模法则的统一分析,揭示了一种贯穿组织层次的一致的亚线性扩展模式——从网络内的群落到整个生态系统。这种来自不同生态系统类型的,复杂食物网的亚线性捕食者-被捕食者缩放的规律,对于理解跨越大空间梯度的自然系统的能量流具有重要意义。
当然,看到这些普适的现象,我们肯定要问,这些亚线性的规模法则是怎么出现的?到目前为止,我们其实还不知道有什么生态学理论,足以解释具有如此丰富多样性的群落具有的普适规律。但可以做一些合理的假设:在稳定状态下,捕食者生物量由一定量的猎物所维持,这意味着,随着猎物生物量的增加,它们的总产量应该扩大到接近 3/4,以匹配它们所支持的捕食者生物量。因此,依赖密度的过程,例如对资源的竞争和被捕食物种之间的其他消极的相互作用,可能导致平均增长率以亚指数级下降。
当然,现有理论还远远不能给出完整的解释,但本文的研究结果揭示了捕食者和被捕食者的生物量比例在不同食物网内和不同食物网间基本相似性,为连接不同生物组织层次的生物量的分布提供了基础。
陶如意 | 作者
邓一雪 | 编辑
商务合作及投稿转载|swarma@swarma.org◆ ◆ ◆
搜索公众号:集智俱乐部
加入“没有围墙的研究所”
让苹果砸得更猛烈些吧!
,