【科研摘要】
大自然为人们提供了具有从分子构件到层次缔合行为的巨大结构多态性的原纤维全景。尽管最近在通过自组装创建具有单个构件的人造系统方面取得了成就,但是分子编码从模型构件到原纤维缔合的关系(导致受控的宏观特性)的分子编码仍然是一个遥不可及的目标。

最近,复旦大学陈国颂教授团队通过使用一组设计的糖肽结构单元,并结合实验和计算工具,报告了一个受控的原纤维多态性库,阐明了从分子堆积到原纤维缔合以及相关的宏观性质。右旋或左旋扭曲在原纤维的轴向或径向生长取决于寡糖和寡肽成分之间的微妙权衡。同时,实验和理论上提出了双链原纤维缔合过程的可见证据。最后,原纤维多晶型物在水凝胶形成和细胞迁移控制上表现出显着不同的宏观性质。相关论文以题为A Comprehensive Landscape for Fibril Association Behaviors Encoded Synergistically by Saccharides and Peptides发表在《J. Am. Chem. Soc.》上。
【主图导读】
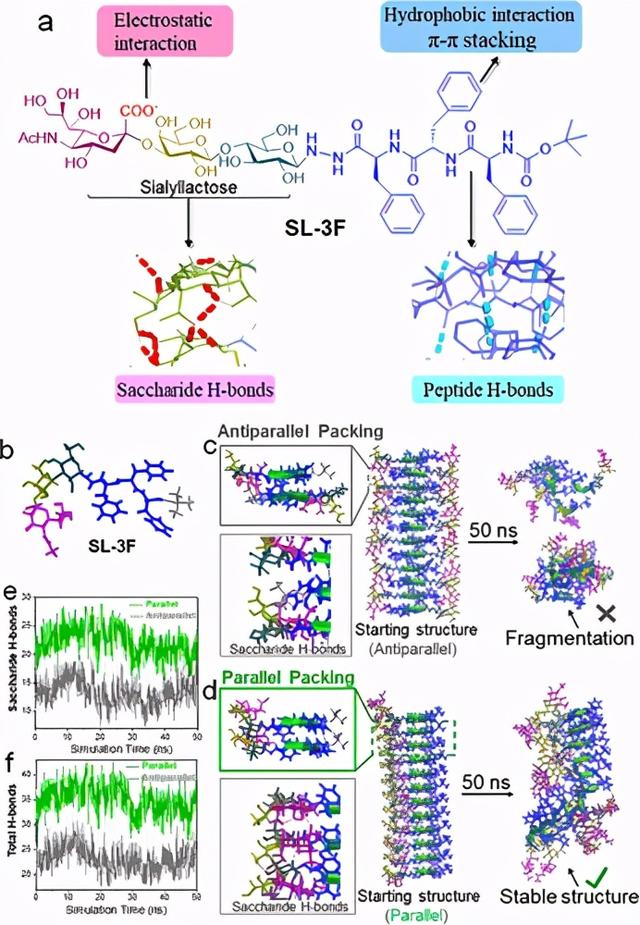
示意图1.(a,b)SL-3F的代表性模型,(c)反平行堆积模式和(d)平行堆积模式的全原子分子动力学模拟结果,由(e)糖化物部分形成的氢键的定量比较( 糖H键)和(f)两种组分(糖部分和肽部分)(总H键)处于平行包装模式和反平行包装模式(12×SL-3F)

图1. SL-3F的原纤维缔合行为(1 mg/mL的SL-3F在水中)。(a,b,d,e)AFM高度图像和双股原纤维的相应轮廓,(c)负染色TEM图像,(f)SL-3F轮廓长度的统计结果,(g)显示长宽比的图 与各种纤维组件的结构单元分子量的关系

图2. SL-3F的缔合过程与孵育时间的关系;所有比例尺均为50 nm。

图3.双链工艺的全原子模拟。(a)采用平行堆积方式的SL-3F单链原纤维的松弛过程。(b)在糖部分模糊之后,SL-3F的单链原纤维的顶视图和正视图。(c)SL-3F的每个苯丙氨酸残基的溶剂可及表面积(SASA)的平均统计结果,(d)说明链形成和链缔合之间的竞争。(e)SL-3F的双链原纤维的松弛过程。(f)苯丙氨酸残基的SASA值在经历双链过程之前和之后有助于链缔合,(g)涉及SL-3F双链原纤维的两种H键的统计结果,(h) SL-3F双链原纤维的CD结果(1 mg/mL,水)。

图4. SL-4F的原纤维缔合行为。(a,b)TEM的负染色(c)SL-4F的扭曲带状结构的冷冻EM图像,(d)糖部分模糊后SL-4F的单链原纤维的俯视图和正视图,(e)SL-4F的每个苯丙氨酸残基的溶剂可及表面积(SASA)的统计结果,(f)链形成和链缔合竞争的图解,(g)径向相关原纤维的AFM高度图SL-4F,(h)SL-4F扭曲带的松弛过程(正视图和侧视图),(i)与SL-4F的原纤维有关的两种H键的统计结果,(j)CD SL-4F的结果; a,c和g中的比例尺分别为50、50和10 nm。
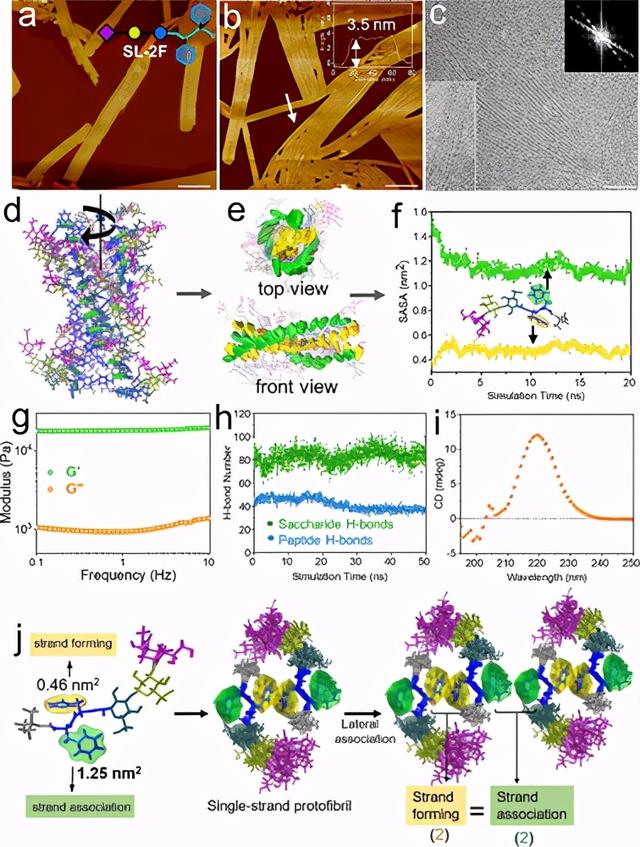
图5. SL-2F的原纤维缔合行为。(a,b)AFM高度图像,(c)SL-2F多股带状结构的Cryo-EM图像,(d)单股原纤维的全原子模拟结果,(e)单股原纤维的顶视图和正视图 糖部分模糊后,SL-2F的原链原纤维,(f)SL-2F的每个苯丙氨酸残基的溶剂可及表面积(SASA)的统计结果,(g)SL-2F凝胶的流变行为(0.1质量% ),(h)SL-2F的原纤维中涉及的两种H键的统计结果,(i)SL-2F的CD结果,(j)链之间竞争的图解说明- 形成和链缔合;a,b和c中的比例尺为100 nm。

图6.(a)涉及原纤维的两组H键的统计结果,以及(b)不同糖肽的流变行为(0.5质量%)。由模拟时间内的平均糖H键计算出的糖H键/肽H键(S/P)除以模拟时间内的平均肽H键。(c)水凝胶强度(G')随糖H键与肽H键的数量比(S/P)的变化而变化。

图7.(a)Transwell分析的示意图,(b)各种原纤维对B16F10细胞迁移的体外抑制作用,(c)SL-3F双链原纤维对B16F10细胞生长的体外抑制,可见 (d)CLSM和(e)FE-SEM分析SL-3F双链原纤维与B16F10细胞之间的相互作用。
【总结】
团队已经在实验和理论上基于一组设计的糖肽模型论证了糖和肽对原纤维缔合行为的协同编码。事实证明,寡糖和寡肽成分之间的微妙权衡是决定生长方向以及原纤维手性的决定因素。原纤维缔合也被认为是在水凝胶化和抗肿瘤活性方面桥接分子构件和一些宏观性质的有用工具。值得注意的是,发现具有高达105的超高纵横比的SL-3F的双链原纤维能够在溶液中高度存活并牢固地粘附在肿瘤细胞表面上。糖基化是自然界中普遍存在的现象,它参与各种具有不同作用的纤维缔合过程。作者希望,糖在纤维缔合上的分子细节可能揭示出聚糖参与的一些机制,并预测未阐明的学科。简而言之,所报道的原纤维缔合行为的全面概况可能提供工具,以解码天然存在的原纤维缔合并在纤维材料设计上获得独到的见解。

TOC.
参考文献:doi.org/10.1021/jacs.1c01951
版权声明:「高分子材料科学」是由专业博士(后)创办的公众号,旨在分享学习交流高分子聚合物材料学等领域的研究进展。上述仅代表作者个人观点。如有侵权或引文不当请联系作者修正。商业转载或投稿请后台联系编辑。感谢各位关注!
,




