基金项目:柑橘果实浮皮的研究与展望
基金项目:国家重点研发计划项目( 2017YFD0202000 );国家现代农业产业技术体系建设专项资金项目( CARSG26 )
莫小荣等
1 柑橘果实浮皮症状和衡量
柑橘“浮皮”是一种在宽皮类柑橘果实成熟后期常发生的生理性病害,主要表现为果皮表面粗糙、油胞大,果皮浮起发泡、粗厚,海绵层组织松散、结构软弱、排列 无序甚至塌 陷,海绵层和 外果皮连接 疏松 ,果皮与囊瓣膜隔层变大甚至分离 ;温州蜜柑浮皮果果皮和果肉之间的空隙约占整果横切面宽度1 / 20 ;与不易发生浮皮的杂柑类、橙类、柚类相比,易浮皮的宽皮柑橘白皮层组织浮皮程度高,细胞密度低,细胞结构松散 .如图 1 所示,柑橘浮皮程度可分为 4 个等级,0 级:果面无浮皮; 1 级:轻度浮皮,浮皮斑 ≤1 / 3 果面;2 级:中度浮皮, 1 / 3 果面 < 浮皮斑 <2 / 3 果 面; 3 级:重 度 浮 皮,浮 皮 斑 ≥2 / 3 果面 .浮皮程度用浮皮指数即[Σ (浮皮级果数 ×代表级值)/(调查总果数 × 最高浮皮等级代表值)]表示,还 可 以 采 用 浮 皮 空 间 率 和 比 重 测 定 法 来衡量 .
2 柑橘浮皮果实的品质特征
浮皮果多呈粗皮大果,果实质量降低且易受伤

图 1 浮皮程度分级
腐烂,果实耐贮运性下降 .浮皮果常伴有果皮表面凹凸不平、色泽泛黄甚至发生异臭,囊瓣膜增厚、咀嚼性差,汁胞失水、干硬,营养物质缺乏,果实整体风味寡淡,食之无味,甚至失去食用价值 .浮皮果有机酸、糖和维生素 C ( vitaminC , Vc )等均明显减少,尤其是可溶性糖和有机酸 ;随着南丰蜜橘果实浮皮程度加重,果实可溶性固形物(totalsolublesolid , TSS )、可滴定酸( totalacid , TA )和 Vc含量迅速降低,果实浮皮与 TSS 、 TA 和 Vc 呈负相关 ;胁山特早熟温州蜜柑果实在出现轻微浮皮症状后果皮率增加、可溶性糖含量下降 . Vc 含量下降 是 果 实 衰 老 的 表 现,果 实 浮 皮 表 明 其 已 衰老 .浮皮果糖酸含量下降,有人认为是浮皮果呼吸作用强而导致可溶性糖、有机酸类物质被大量消耗 ,也有人认为是浮皮果果肉营养物质和水分通过果皮与果肉间维管束(橘络)传输给果皮而诱发果皮二次生长 ,还有学者认为是果实聚合多糖转化为可溶性碳水化合物的过程受阻所致 .相反,浮皮果棉子糖含量不降反升 .
3 柑橘果实浮皮发生的条件
3.1 果实大小、成熟期与浮皮发生
果实浮皮主要发生在温州蜜柑、椪柑、沙糖橘等易剥皮的宽皮类柑橘,而果皮硬、紧实难剥的甜橙、柠檬等柑橘则不易发生.浮皮果主要发生于果实成熟后期 ,但特早熟温州蜜柑在果实成熟期前已经出现轻微浮皮 .细胞学动态变化特征分析结果显示浮皮发生的关键时期为花后 85~110d ,而花后 7~28d 是粗皮大果的温州蜜柑果皮增厚的关键时期 .四川红桔浮皮率在 12 月中旬、1 月中旬、2 月中旬分别达到 25% 、 85% 和 100% ;沙糖橘横径 M 级( 45~50mm )果浮皮率在 12 月中旬、1 月中 旬、 2 月 中 旬 分 别 为 42.22% 、 55.56% 和100% ,而延时采收容易因果皮老化而出现浮皮加重,采收越迟,浮皮率越高、浮皮程度越重 .近年来,为了提高温州蜜柑、椪柑等宽皮柑橘果实糖度,很多地区推出完熟栽培技术,增加了果实 TSS ,但也提高了果实浮皮的发生率,降低果皮硬度和贮藏性 .强旺树、幼树、小年树果实及朝天果、顶果、大 果、粗 柄 果 容 易 发 生 浮 皮,形 成 粗 皮 大果 ,早熟温州蜜柑果实浮皮程度随果实增大而加重 .沙糖橘横径 2L (55~60mm )、 L ( 50~55mm )、 M ( 45~50mm )、 S ( 40~45mm )和 2S ( 35~40mm )果 实 的 浮 皮 指 数 依 次 为 91.67 、 43.70 、21. 11 、 5. 56 和 0 ,果实越大,浮皮发生时期越早、程度越重,浮皮率、浮皮指数越高 ,因此,生产上要多保留中、小形果实而尽早疏除 2L 级以上大果.
3.2 矿质营养对浮皮发生的影响
不科学施用壮果肥会加重果实浮皮程度,延迟施用壮果肥会导致树体氮过多而促进果实增大,转色延迟,助长果皮二次生长,易诱发浮皮 .钙作为植物细胞的结构物质,能稳定细胞壁和细胞膜结构,决定着果实硬度,果实膨大期钙不足会导致果皮细胞排列松散、韧性降低,易发生浮皮 ;钙在细胞壁的分布与温州蜜柑内果皮细胞壁的生长或变形密切相关,并直接影响浮皮的形成 ,而红桔浮皮果果皮钙的含量明显降低并与浮皮现象呈显著负相关 ;浮皮果实经钙化合物处理后细胞间隙变小,细胞壁纤维物质致密,细胞壁肿胀少,未发生细胞分离 ,因此,喷施钙肥可改善果皮物理性质,提高果皮钙含量和果实品质,一定程度上能抑制浮皮的发生 .柑橘缺硼也容易造成树体缺钙从而诱发浮皮 .浮皮果常伴随果实枯水和粒化,Mosambi和红血甜橙粒化过程中叶片锌和铜含量升高 ,粒化琯溪蜜柚果肉有较高的铜锌含量 ,锌和铜在柑橘浮皮发生过程中也起到一定作用.
3.3 气候因素对浮皮发生的影响
新鲜果蔬作物的产量和质量直接或间接地受高温的影响,温度升高直接影响植物光合作用,引起糖、有机酸、类黄酮含量,硬度和抗氧化活性的改变 .温州蜜柑从花后至生理落果期及成熟期的温度升高 2℃ 时显著增加果实浮皮率,降低果实TSS 和 TA 含量 .柑橘壮果期遇到干旱、降雨少,或在果实转色期和成熟期遇到大雨、高温高湿天气、土壤湿度大均容易发生浮皮 ;克里曼丁和椪柑采收后果实浮皮加重且含水量显著增加 ,为避免果实枯水而采用高湿环境贮藏温州蜜柑、椪柑等宽皮柑橘加重了果实浮皮的发生,贮藏过程发现果肉通过发达的维管束系统将大量水分运输至果皮,导致果皮吸水膨大,果皮和囊瓣膜之间产生空隙而加剧了果皮浮皮 .
3.4 其他因素对浮皮发生的影响
高 CO 2 和低 O 2 的贮藏条件下温州蜜柑果实浮皮率显著提高和浮皮程度加深 ,这可能与果实发生无氧呼吸有关.病虫危害下,柑橘果实会发生应激反应从而进行自我保护,果皮细胞迅速分裂和增大,从而形成浮皮果 .
4 柑橘果实浮皮发生的机制
4.1 细胞形态与结构变化
柑橘果实浮皮主要是指果皮和囊瓣膜间隙增大的现象,空隙变大的原因可能是由于果皮细胞胞间层的崩解而导致的细胞间连接松散及细胞壁肿胀所致 .浮皮形成过程中,油胞层细胞分裂时间最长,细胞数量增多和体积增大较快;海绵层外侧细胞次之;海绵层内侧最短,海绵层受油胞层牵制和拉扯,最终出现海绵层与油胞层空隙扩大、疏松甚至断裂,也就是果皮不同部位生长发育不同步造成浮皮 .花后 85d 浮皮果的海绵层细胞出现轻微疏松、边缘细胞分离、细胞间隙变大,而靠近油胞层周边细胞则排列整齐紧密;反之,耐浮皮的芽变果海绵层细胞排列紧密,直至果实发育成熟才开始出现较大间隙,由此推测果实浮皮形成过程和细胞生长发育密切相关,主要由海绵层和油胞层生长发育不同步造成 .张世怡 发现花后 88~113d 宫川温州蜜柑果实内果皮处的薄壁细胞较其芽变果排列混乱、细胞间隙大;花后 113d ,白皮层细胞开始出现细胞壁结构疏松,质壁分离,细胞内容物降解,细胞核结构不完整,叶绿体体积增大,基粒结构松散、无序甚至完全降解,类囊体衰退,叶绿体中部出现嗜饿体或油体等;花后 165d ,白皮层细胞壁解体、原生质体降解、质壁完全分离及叶绿体降解等,其芽变果细胞膜呈现轻微质壁分离而细胞核结构相对完整.
4.2 酶活性变化
柑橘果实浮皮与果皮代谢活动密切相关.如红桔浮皮果的果皮原果胶含量显著低于正常果,主要是因为浮皮果的果胶甲酯酶(pectin methylesterGase , PME )活性高,导致浮皮果果胶物质分解、细胞壁降解,PME 活性升高与果实衰老有关.果皮中多聚半乳糖醛酸酶(polygalacturonase , PG )活性和丙二醛( malondialdehyde ,MDA )含量在特早熟温州蜜柑胁山浮皮形成过程中不断上升,超氧化物歧化酶(superoxidedismutase , SOD )活性不断下降,表明浮皮果果皮活性氧清除能力下降、膜脂过氧化作用加强、果皮衰老加剧 .温州蜜柑浮皮程度加重过程中也伴随着果皮中原果胶和可溶性果胶含量下降、 MDA 积累以及 PG 、纤维素酶(cellulase , Cx )等细胞壁水解酶活性升高,即果皮浮皮形成过程也是果皮膜脂过氧化作用加强的衰老过程 .宫川浮皮果果皮 SOD 活性在果实发育初期较高,随果实成熟而不断下降,耐浮皮的芽变果 SOD 活性高于宫川,表明自由基清除系统失调 .胡柚果实贮藏中、后期果肉 SOD 活性下降与果皮 SOD 活性上升同步,即果肉组织衰老和果皮组织再生长同步是果实枯水(浮皮常伴随果实枯水和粒化)发生的根本原因 .喷钙使南丰蜜橘果皮过氧化氢酶(catalase ,CAT )、 SOD 活性提高而 MDA 含量和过氧化物酶(peroxidase , POD )、多酚 氧 化 酶 (polyphenoloxiGdase , PPO )、 PG 、 PME 、 Cx 等细胞壁水解酶活性降低,对防治果实浮皮具有一定作用 .
4.3 内源激素变化
柑橘果实浮皮的发生过程也受内源激素调控.乙烯(ethylene , ET )会加剧浮皮的发生,而吲唑酯的代谢物 5G 氯 G3G 吲唑甲酸能够抑制 ET 的合成,在果实着色前期喷施吲唑酯可以减少浮皮的发生 .宫川浮皮果白皮层 IAA 、 ABA 含量均高于芽变果,但果实成熟后期浮皮果白皮层 ABA 急剧升高,而芽变果则快速下降,说明果实成熟后期浮皮果白皮层衰老因果皮生长撕拉形成浮皮 ;克里曼丁和椪柑浮皮果果皮 ABA 大量积累且均显著下调,与 β G胡萝卜素合成降低及 ABA 分解加快有关,浮皮的发生可能是果皮衰老延迟所致 ;本地早柑桔果实贮藏期间果皮 ABA 和 IAA 积累增加,果肉 ABA含量却不断降低,以此推测果肉组织衰老先于果皮,果皮出现二次生长 .因此,浮皮的产生源于白皮层和果肉先衰老、果皮延后衰老并发生二次生长.喷施外源赤霉素(gibberellinA 3 , GA 3 )可以降低外果皮 ABA 含量 ,增加白皮层的致密性而改善果实浮皮现象 ;特早熟温州蜜柑胁山在浮皮至最终汁胞粒化过程中果皮和果肉组织内源多胺(polyGamine , PA )总量下降、[精胺( spermine , Spm ) + 亚精胺(spermidine , Spd )]/腐胺( putrescine , Put )比值不断下降,且果皮先于果肉,喷施外源 Spd 可显著增加内源 PA 总含量,提高( Spm +Spd )/ Put 比值,降低浮皮的发生 .黄果柑果实浮皮过程中(Spm + Spd )/ Put 比值降低,抗氧化 酶 ( SOD 、CAT )活性降低,而加速果实衰老以及浮皮粒化 ,而外源喷施 Spd 也降低黄果柑果实枯水(浮皮) .此外,外源添加 GA 3 处理,能提高早熟温州蜜柑果皮 SOD 活性,降低 PG 活性和 MDA 含量,减少浮皮的发生 .
4.4 基因表达与代谢途径变化
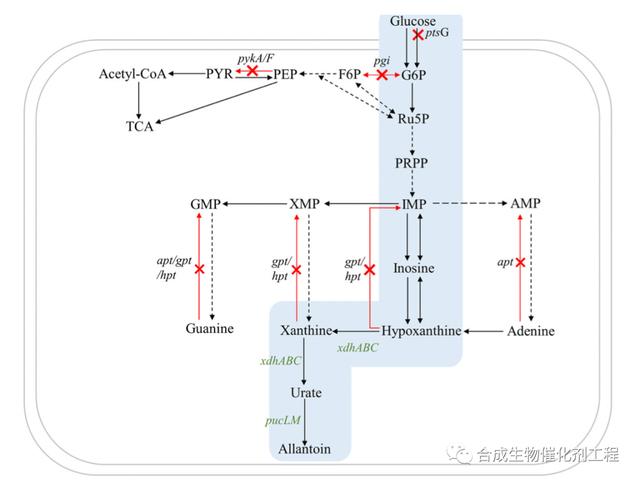
对不同柑橘品种的正常果与浮皮果不同部位差异表达基因的分析结果表明,浮皮果白皮层大部分CTK 、 GA 3 信号传导基因表达下调,糖类转化相关基因表达上调,推测白皮层是浮皮发生的主要部位,浮皮的发生与果实成熟早期参与初始代谢基因表达差异密切相关 ;在白皮层发育阶段,宫川芽变果白皮层的赤霉素信号刺激调控基因( GAGstimulatedinarabidopsis4 , GASA 4 )和细胞分裂素受体蛋白基因(cytokinindependent1 , CKIG 1 )表达量高于浮皮果,表明幼果时期芽变果赤霉素和细胞分裂素代谢旺盛,而浮皮果则处于较低水平 ;张世怡 对浮皮形成关键时期的温州蜜柑宫川及其芽变果白皮层转录组进行分析,获得 733 个差异表达基因,与膜组织、脂类相关合成与代谢、氧化还原酶及催化酶活性等有关.蔗糖、淀粉代谢途径以及甘油酯、脂肪酸、戊糖磷酸等初级代谢途径抑制赤霉素信号刺激调控基因( GAGstimulatedinarabidopsis1 ,GASA 1 ;GAGstimulatedinarabidopsis 5 , GASA 5 )表达;一些 热 激 蛋 白 ( heat shock proteins , HSPs )如HSP70 、 HSP82 及热激因子 A4 表现为浮皮的主要转录调控枢纽 ;浮皮果果皮衰老与ABA 在果皮的积累密切相关, ABA 的积累与 ABA 生物合成途径中 β G 胡萝卜素含量下降和番茄红素 β 环化酶基因(lycopene β Gcyclase1 , LCYB 1 )、番茄红素 ε 环化酶基因(lycopeneεGcyclase , LCYE )、脱落酸 8′G 羟化酶基因(abscisicacid8′Ghydoxylase , AB 1 )的表达密切相关 .姚世响等发现椪柑浮皮果果皮挥发性物质组分未发生变化,但总含量降低了 82% ,其中占比最大的萜烯类物含量降低了 81.4% ,且萜烯类物质合成基因表达量明显下降,浮皮果挥发性物质总量下降与萜烯类物合成途径受抑制密切相关.由此推测,柑橘果实浮皮发生的分子机制涉及赤霉素和细胞分裂素信号传导、萜烯类物质合成基因表达下调,影响初级代谢及膜脂、蔗糖和淀粉的合成.柑橘果实浮皮形成过程及机制如图 2 所示.柑橘果实浮皮表现为果皮和囊瓣膜间隙异常增大,一方面是由于果肉糖酸、水分不足,容易出现果肉枯水、干瘪,增加果皮和囊瓣膜的间隙;另一方面是果皮结构异常,果皮细胞间连接松散、细胞壁肿胀从而增加果皮和囊瓣膜间隙.其中果皮结构异常是关键,其过程大致为:外界作用因子如水分、气体、养分、生物及干预等导致果实内源激素( ET 、 ABA 、IAA 、 CTK 等)失调,进而扰乱果实代谢相关酶( Cx 、PG 、 PME 等)活性,果实内源激素和代谢相关酶活性同时受基因调节,引起果皮原果胶降解、膜脂过氧化,出现海绵层细胞壁解体、质壁分离,诱发海绵层、油胞层再生发育不同步,形成海绵层松散、间隙大,油胞层整齐、紧密,加剧果皮与囊瓣膜间隙增大,最终形成了浮皮。

图 2 柑橘果实浮皮形成过程及机制
5 展 望
柑橘果实浮皮主要表现为果皮浮起、增厚、凹凸不平、细胞间隙大,海绵层软化、崩溃,果皮与囊瓣膜间隙大;浮皮果果皮粗厚、易腐烂,果肉糖、酸含量低,囊瓣膜厚硬、汁胞失水硬化.柑橘果实浮皮发生始于花后约 3 个月即海绵层发生细胞壁结构破坏,随着果实成熟发展到质壁分离、原生质降解及细胞器衰老;果实偏大、成熟期高温高湿、晚熟品种以及过量施用氮肥、延迟施用壮果肥、硼和钙营养失调等均诱发或加剧浮皮发生.已有研究表明,柑橘果皮不同部位发育不同步及果皮海绵层提前、加速衰老是浮皮发生的主要机制,涉及果皮赤霉素和细胞分裂素信号传导及萜烯类物质合成基因表达下调、内源激素失调、抗氧化酶活性降低而细胞壁水解酶活性增加等过程,意味着柑橘果实浮皮的发生和形成是 一 系 列 复 杂 的 生 理 生 化 作 用 的 结果 .简单而言,柑橘果实浮皮,一是果皮和囊瓣膜间隙增大,二是海绵层空隙扩大,即果皮不同部位生长发育不同步,因此,“不同步发育”是关键,尤其是阻止白皮层细胞壁解体、质壁分离.基于此,柑橘果实浮皮亟需深入研究 3 个层面的问题:一是“源库供应”上,需要研究揭示“源(叶)”、“库(果)”光合产物、营养元素传输、分配对浮皮发生、形成的调节作用与机制;二是“竞争抑制”上,需要研究揭示新梢、叶片、果实间碳、氮等代谢及养分分配的相互制约和联系,明确水、肥、气、热“靶向调节”的作用和机制;三是“整体协调”上,需要研究揭示不同品种、不同区域柑橘根系活动、新梢(叶片)生长、花芽分化、果实发育的相互关系,明确物候期的生长中心和调节要领,通过“整体协调”与“靶向调节”破解柑橘果实浮皮难题。
,