支链淀粉的中长链(12≤DP≤24)含量影响淀粉颗粒的晶化
研究认为,支链淀粉结构对淀粉颗粒的形态及物理化学性质有着直接的影响,其中支链淀粉的中长链对支链淀粉尤其重要。(结构决定物理化学性质)
支链淀粉的中长链含量能够影响淀粉颗粒的晶化。研究表明,支链淀粉的短链 (DP<11) 太短, 不能形成跨越整个结晶片层的双螺旋结构, 会导致结晶区域缺陷和晶体结构的不稳固。另一方面, 中长链 (例如DP 16~21) 却能够形成跨域整个结晶片层的双螺旋结构,有利于淀粉颗粒的结晶[1]。淀粉颗粒的结晶程度越高,淀粉颗粒就会越致密,反之,结晶程度越低,淀粉颗粒就会变得越来越疏松,直至变成可溶性淀粉。糖化胚乳(sugary)突变体中可溶性淀粉和高度分支化的多糖(植物性糖原)含量极高,而野生型种子中可溶性淀粉含量几乎没有,与此同时sugary突变体中淀粉含量显著减少,种子皱缩[2, 3]。其中sugary-1突变体表型最为明显,与之相对应的是sug-1的短链(DP<12)含量极其显著增加,高达58.3%,而中长链(12≤DP≤24)含量显著降低[4]。

淀粉颗粒的结构模型:中心点为原点,是淀粉合成起始的地方。支链淀粉相邻分支之间是双螺旋结构,6个葡萄糖分子为一个螺旋。支链淀粉内的双螺旋的排列结构决定了他们的晶体构型,A型大部分为谷物淀粉,B型大多为马铃薯等非谷物的淀粉
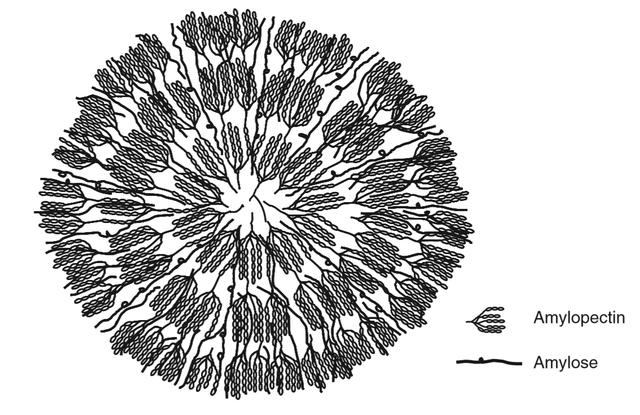
淀粉颗粒的二维结构图:双螺旋集中的一层为半结晶层,是淀粉具有晶体性质的原因

大部分半晶体层厚度为9-10nm,而他对应的最长的链长约为32
支链淀粉的中长链(12≤DP≤24)含量影响淀粉颗粒的晶化程度进而影响种子充实程度
淀粉的晶化程度对淀粉颗粒的致密程度有着重要的作用,进而会影响种子的充实程度,这个是毋庸质疑的。也就是说当支链淀粉的中长链(12≤DP≤24)含量少到一定程度(程度不同,所呈现的表型也不同)是会影响种子的充实程度,也可以换一另一种方式来解释:当支链淀粉的短链(DP<12)的含量过高时会影响种子的充实程度。
淀粉合成相关酶的突变导致的种子干瘪主要有三类酶,shr1和shr2突变体(AGPase)、sug-1突变体(ISA1)、Pho1的突变体,其他淀粉合成相关酶的突变只会导致种子垩白增加,或者改变淀粉的含量[5]。而这三类突变体的支链淀粉链长变化有一个共同特点,DP 6-12的短链增加,而DP 13-24的中长链减少[5]。这个证据足以表明支链淀粉的中长链对种子的充实程度有着重要的意义。
支链淀粉的中长链(12≤DP≤24)含量影响水稻种子的籼粳分化
水稻分为籼稻和粳稻,他们之间的支链淀粉链长分布有所区别,原因可能是籼稻和粳稻之间SS1/SS2a的活性区别所导致的,研究发现粳稻中的SS1活性高于籼稻而SS2a活性远低于籼稻,导致籼稻短链(DP<12)的含量较低而中长链(12≤DP≤24)的链长含量较高。以∑DP≤11/∑DP≤24的链长比值作为划分标准,比值小于0.22的为L型,比值大于0.26的为S型,大部分籼稻都为L型,少部分为中间型,基本上所有的粳稻都为S型[6, 7]。
彭小松等利用重组自交系研究表明, 群体中支链淀粉短链 (6≤DP<12) 的含量逐渐增高(中长链含量逐渐降低)表现为籼型<偏籼型<偏粳型<粳型(籼粳杂种后代支链淀粉结构及其与稻米糊化特性相关性分析)。而随着短链含量的不断增加,由正常粳稻的短链含量(35%-38%)增加到sug-1突变体的短链含量(58.3%)[4],种子的表型也就越有可能趋向于糖化胚乳(sugary)的表型。在玉米、水稻、大麦、马铃薯当中都存在sugary突变体,最严重的就是ISA1的突变体(sug-1),而玉米中存在糖化胚乳表型较弱的突变体是SS2a的突变所导致的[5]。在水稻中找到了另外一种表型较弱的糖化突变体sug-2,它的表型为胚乳中心糖化,不能被碘液染色,研究认为sug-2调控了PUL(普鲁兰酶,另外一种DBE)的表达。Fujita等将PUL突变后,发现短链的含量显著增加,但变化程度远小于sug-1[8]。这说明PUL跟ISA1一样也参与了支链淀粉短链的酶解, PUL功能与ISA1部分重叠,并且其缺陷对支链淀粉的合成的影响比ISA1缺陷小得多(PUL)。那么,粳稻相对于籼稻缺少了SS2a这个合成中长链的酶的活性会不会导致一定的糖化表型呢?我想一定是会的。在半薄切片碘液染色观察中,发现了有相当一部分ZH11(一种粳稻)种子中心区域并不能被碘液染上颜色,同时中心区域的淀粉颗粒非常小且边界并不明显,这很有可能就是粳稻胚乳糖化的表现。

图为本人拍摄的水稻种子半薄切片碘液染色种子中心区域,可以明显看到染色的深浅有不同,染色浅的区域即为糖化区域
假设将糖化程度的不同和支链淀粉的中长链(12≤DP≤24)含量联系起来,那么随着支链淀粉的中长链 (12≤DP≤24)含量逐渐降低,种子表现为籼型<偏籼型<偏粳型<粳型<中心胚乳糖化<胚乳轻微糖化<胚乳重度糖化。其中sug-2属于中心胚乳糖化,而sug-h(ISA1和BE2a的双突变体)属于胚乳轻微糖化,sug-1属于胚乳重度糖化。糖化的程度可以用碘液染色的程度以及半薄切片观察来区分,糖化程度越高,胚乳充实和淀粉积累程度越低。
支链淀粉的中长链(12≤DP≤24)合成模型
前人的支链淀粉链长合成的研究及模型明确了BE、SS、ISA等各种酶之间的分工。

但很多研究都表明,支链淀粉合成相关酶之间的关系并不是明确割裂的,而是你中有我的合作关系[9]。现有的模型是将有特定功能的酶类区分开来,合成短链的基因是SS1,合成中长链的基因是SS2a,合成长链的基因是SS3a。但是这个模型所涉及的酶类还并不完整,且有时候不能解释某种酶突变以后造成的支链淀粉链长的变化。如Pho1突变后短链增加,中长链减少。AGPase的突变也会导致短链增加,中长链的减少[5]。同时某些基因例如SS1,SS2a,BE2a,PUL这些酶突变以后为什么不会有显著的可以观察到的表型,这些问题都需要我们去回答。
那么现在我们假设支链淀粉合成过程中,植物为了更多地储藏淀粉,它们将趋向于合成中长链(12≤DP≤24)或者去除短链(DP<12)以利于淀粉颗粒的结晶。由此,将植物淀粉合成酶分为四类,一类是用来合成中长链的,一类是用来去除短链的,一类是合成长链和超长链的(这一类先不做讨论),最后一类是合成短链的。
哪些酶能够合成中长链呢?首先是SS2a,活性测定发现,它倾向于延伸分支点外侧的短链。Nakamura等将籼稻IR36的SSIIa的cDNA引入到粳稻品种中,会使DP 10的支链淀粉转化为DP 12-24的支链淀粉[10]。由此,SS2a被认为是合成中长链的主要酶类。然后是BE1,支链淀粉链长分布变化发现BE1的突变体中DP 11-22的链显著降低而DP 23-36的链增加,通过活性测定发现BE1通过支化较少支化的葡聚糖优先产生较长DP>=16的支链[11]。虽然BE1的活性不是延伸链长,但是它特异的分支酶活性却可以直接形成中长链。由此,我们也可以将BE1划为中长链的合成酶类。最后是Pho1,支链淀粉链长分布变化发现Pho1的突变体中DP<12的短链显著增加中长链DP 12-24含量显著降低[12]。先前的酶动力学实验认为,Pho1主要在淀粉代谢中起着降解的作用,然而,最近的酶动力学实验认为它在淀粉的生物合成中起着重要作用。将游离的磷酸基团(Pho1合成淀粉方向的产物)加入进去,淀粉的合成仅受部分影响,而不是反过去分解淀粉。因此,他们认为虽然Pho1处于有利于淀粉降解的条件下,Pho1优先进行生物合成反应[12]。在体外利用重组酶(rPho1和rSSIIa)对DP=6的MOS延伸,发现Pho1延伸MOS的能力要比SSIIa要高[12]。虽然Pho1在体外实验中被证明对淀粉的延伸没有偏好性,但是它在体内却能够将短链延伸成中长链,或许在体内存在有特殊的机制可以让Pho1来合成中长链,因此我们仍将Pho1划为中长链的合成酶类。
哪些酶能够去除短链呢?显而易见,解分支酶DBE是去除短链的最主要方式。其中ISA1是分支酶活性最主要的成分,当ISA1突变后,短链含量会增加到60%左右。PUL(普鲁兰酶)也是解支链淀粉酶的一种,将PUL突变后,支链淀粉的短链含量显著增加,但变化程度远小于ISA1的突变体。
哪些酶能够合成短链呢?首先是SS1,支链淀粉链长分布变化发现SS1的突变体DP 8-12的支链淀粉大量减少而DP 6-7的支链淀粉大量积累,体外酶活也证明SSI优先合成短链(DP 6-15)。SS1的活性约占总SS活性的70%,由此,SS1被认为是合成短链的主要酶类。其次是BE2a和BE2b,BE2a产生6-11的短链,而BE2b产生6-7的短链。
按照假设,合成短链的酶对淀粉的结晶是存在负效应的,它们的酶活性与前面的那两类合成中长链和去除短链的酶的活性是拮抗的。是否有研究能够证明这个假设呢?答案是肯定的。有研究将拟南芥中合成淀粉的酶类按照不同的组合转入酵母体内,在酵母体内合成了淀粉颗粒,发现合成短链的SS1对不溶性的淀粉(或者葡聚糖)的含量有极其显著的负效应。另外,在水稻中发现了一个较弱的糖化突变体sug-h,它是ISA1和BE2a的双突变体,也就是说突变BE2a能够减弱ISA1突变所造成的胚乳糖化。将sug-h中BE2a超量表达以后,sug-h的表型又变成了较为严重的sug-1的表型。
接上所述,既然合成短链对种子淀粉颗粒结晶是不利的,那么这几个酶在淀粉合成过程中又有什么存在价值呢?存在即合理。它们合成出来的短链对支链淀粉的结晶是不利的,那么肯定存在有能够延伸这些短链的酶。其中一个是SS2a,但是在水稻中又存在着一个问题,粳稻中的SS2a活性远远低于籼稻,在缺乏SS2a活性的情况下,粳稻是如何合成足够多的中长链的呢?由此猜想另外一个能够弥补SS2a活性的酶应该是Pho1。水稻种子淀粉合成酶复合体的共分离实验发现,BE2a和BE2b是能够与Pho1共分离的。但是没有直接证据表明BE2a和BE2b分别与Pho1互作。也许存在有一个特殊的因子能够将BE和Pho1紧密连接起来。
以上:本文主要提出支链淀粉中长链对水稻种子糖化的有决定性的作用,是一个根据前人的研究结果做出的一个合理的假设。同时本文也提出支链淀粉中长链有可能是水稻籼粳分化的一个重要因素,提出粳稻的中心糖化和种子中心垩白是相对于籼稻的一种表型,这种表型是由于中长链的含量变化引起的。更进一步的猜测甚至认为中长链的含量能够影响水稻高温耐受性。这些猜想都需要有实验来证明,我提出这些假设主要希望研究水稻的同学能够与我充分讨论,而不是该专业的同学也能够找出文章存在的逻辑漏洞来讨论。
,




