真菌界概述(二)

真菌学是生物学的一个分支,研究真菌的遗传、生化性质、分类、对人类的用途(包括药用、食用与宗教致幻剂)以及对人类的威胁(包括毒性与感染)。因为许多植物病原是真菌,植物病理学的研究也常与真菌学高度相关。
人类对真菌的利用可以追溯到史前时代,早在5300年前的新石器时代,在奥地利阿尔卑斯山区发现的木乃伊冰人奥茨遗骸上便带有两种多孔菌,其中木蹄层孔菌可能是用来生火的燃料,白桦菌菇则可能是药用。古代人类常在不知情的状况下以真菌为食物来源,例如发酵面包与果汁的生产都有真菌参与。也有早期的手写记录描述作物遭到疑似为真菌的病原感染而破坏。
真菌学是一门相对较新的科学,在十七世纪显微镜发明后才出现。早在1588年,吉安巴蒂斯塔·德拉·波尔塔就发现了真菌的孢子,但真菌学真正的滥觞一般认为是1729年意大利植物学家皮耶尔·安东尼奥·米凯利(英语:Pier Antonio Micheli)出版的《植物新属(英语:Nova plantarum genera)》,他除了观察到孢子之外,还发现在适当条件下孢子可以萌发,并生长成当初产生该孢子的同一种真菌。荷兰科学家克里斯蒂安·亨德里克·珀森被认为是现代真菌学的开创者,他以林奈的二名法为基础建立了第一套菇类分类基础。随后伊利阿斯·马格努斯·弗里斯进一步采集了大量真菌标本,并以孢子颜色与其他微观结构对真菌进行分类,许多标准至今仍被分类学家采用。其他重要的真菌学者还包括迈尔士·约瑟·贝克莱(英语:Miles Joseph Berkeley)、奥古斯特·卡尔·约瑟夫·科尔达(英语:August Carl Joseph Corda)、汉瑞其‧安东‧狄伯瑞(英语:Heinrich Anton de Bary)、路易·勒内·图拉斯内(英语:Edmond Tulasne)与查尔斯·图拉斯内(英语:Charles Tulasne)兄弟、亚瑟·亨利·雷金纳德·布勒(英语:Arthur Henry Reginald Buller)、柯提斯·盖茨·洛依德(英语:Curtis Gates Lloyd)与皮尔·安德烈·萨卡多(英语:Pier Andrea Saccardo)等。二十世纪随着分子生物学、生物化学、遗传学与生物技术的突破,真菌学的研究方法逐渐现代化,DNA定序与支序分类学为真菌多样性与演化的研究打开了另一扇门,也挑战了许多传统以形态为基础的分类系统观点。

许多真菌以菌丝的形式生长,菌丝是直径2-10微米的圆柱体丝状结构,可长达数公分。菌丝一般自其顶端往前生长,新的菌丝形成通常是旧有菌丝的中间产生了一个新的生长点,而长出新的分支,也有些情况是生长中的菌丝顶端分岔成两条菌丝。菌丝成长后相互交错,成为真菌的营养构造菌丝体。菌丝可以是多核(英语:coenocytic)(无隔板(英语:septa))或是有隔板的,有隔板的菌丝不同细胞核间有细胞壁将其分隔,使一个隔间具有一或数个细胞核,多核菌丝则没有隔板,基本上就是一个含有多个细胞核的巨大细胞。隔板上通常有小孔可以使细胞质与胞器流通,有时细胞核也可以通过小孔,例如担子菌门真菌形成的桶孔隔膜。
许多真菌的菌丝形成特化的构造,以更容易从宿主身上吸收养分,例如许多植物病原真菌具有吸器,许多球囊菌门的真菌则形成丛枝(Arbuscular)穿透共生的植物细胞以吸取养份。
虽然真菌属于后鞭毛生物,其共祖有一根后生的鞭毛,但除了壶菌门以及原本属于壶菌门的新美鞭菌门和芽枝霉门外,其他真菌都已经失去了鞭毛。真菌的另一项特征是多数种类具有以几丁质与葡聚糖构成的细胞壁。
(2)宏观结构真菌的菌丝常是肉眼可见的,例如在发霉的墙壁或腐败的食物上,这种菌丝体俗称霉菌,而长在实验室培养基上的菌丝则一般称为菌落,菌落的形状与颜色常是鉴定真菌的重要指标。有些真菌相当长寿,且可以长到非常大的体积,美国马卢尔国家森林的一个蜜环菌菌落已生长到超过900公顷,其生长时间推测已超过9000年。
子囊果与担子果分别是子囊菌门与担子菌门真菌的子实体,内含可以产生有性孢子的子实层。
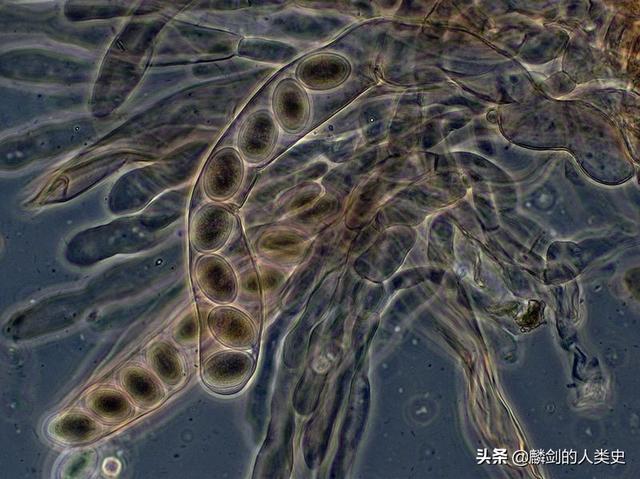
真菌在固体介质中以菌丝形式生长,在水中则常以单细胞酵母菌形式生长,皆是为了提升表面积以增加吸收养分的效率。菌丝特别适合在固体介质上生长,且可以形成较大的穿透力而适于侵入新的固体介质,例如稻热病菌等植物病原菌特化出压器(英语:Appressorium)的构造以穿透植物组织,压器可借由增加细胞中甘油等溶质的浓度以加大膨压,在植物表皮上产生高达1200psi的压力。淡紫拟青霉菌(英语:Paecilomyces lilacinus)也用一种类似的结构来穿透线虫的卵。
绝大多数菌丝都只能自顶端往前生长,Neotyphodium等部分内生真菌(英语:endophyte)则是少见的例外,它们的菌丝附着于宿主细胞上,并可能因宿主细胞延长造成的机械拉力,而促使其菌丝从中间分裂生长(intercalary extension),以长至跟宿主的组织一样长度。另外担子菌子实体形成时,菌柄等大型器官的长成是菌丝体积膨大的结果。真菌形成多细胞组织有数种功能,例如子实体是为了产生与散播有性孢子,生物膜则有助于占据基质与进行细胞沟通,多细胞个体的出现在真菌、动物与植物中都是各自独立演化出来的。
真菌传统上被认为是异营生物,只能依赖其他生物固碳形成的有机碳为碳原,演化造成真菌的代谢能力相当多样化,可以分解各式各样的有机物,有些种类甚至可能使用黑色素从伽马射线等辐射中获取能量,目前只知道少数真菌具有以辐射为能源进行生长的能力,背后的生理机制也不甚清楚,不过整个过程可能与植物利用可见光行光合作用固碳类似,但采用辐射能作为能量来源。
7.繁殖真菌的繁殖机制相当复杂,反映了此类生物生活史与遗传组成丰富的多样性。据统计有超过三分之一的真菌物种采用一种以上的方式繁殖,例如无性时期产生无性孢子,有性时期又产生不同种类的有性孢子。不同的环境刺激可以启动特定基因表现,让真菌长出不同的特化结构以产生无性或有性孢子。
(1)无性生殖真菌的无性生殖有通过产生无性孢子与菌丝断裂(英语:mycelial fragmentation)等机制,前者包括以孢子囊、分生孢子囊柄(英语:conidia)等特化的结构产生孢子,后者则是菌丝断裂成为许多片段,每一片段都长成一条新的菌丝。无性生殖可以产生基因上与母体相同、占据同样生态栖位的子代,繁殖效率也较有性生殖迅速,可以在短时间快速拓殖。不完全菌是一个旧时的分类名词,指未观察到有性世代的真菌,现在分类学上已不使用此名词,但仍可以用来泛指缺乏有性世代的真菌。
(2)有性生殖除了球囊菌门与微孢子虫外,其他门的真菌都可通过减数分裂产生配子、进行有性生殖(且基因组分析显示球囊菌门与微孢子虫的真菌应该也能进行有性生殖,只是尚未观察到)。真菌有性生殖的过程与动物、植物相当不同,而不同真菌间特化来进行有性生殖的结构也有差异,是鉴定真菌物种的重要依据,而拿不同真菌进行交配实验,观察两者能不能形成子实体,可以通过生物物种概念(biological species concepts)以两者能不能成功交配、交配产生的子代有没有生殖能力来判断它们是不是同一物种。主要的生物类群早期都是以有性生殖特化的结构与孢子差异来描述、区分的,例如产生子囊的子囊菌门与产生担子的担子菌门。有一些物种还有交配型之分,必须不同交配型的两个体才可以交配(heterothallic),其他物种则可以和所有个体交配,甚至自交产生孢子(homothallic)。
许多真菌的生活史均存在单倍体与二倍体的时期,真菌进行有性生殖时,菌丝的融合形成交错的网络,这个现象称为融合(anastomosis),是有性生殖不可或缺的。许多子囊菌与担子菌接着会进入双核体(英语:Dikaryon)的时期,即菌丝中一个细胞里有两个细胞核,分别来自两个亲代个体,这两个细胞核不会马上进行核聚变(karyogamy),而会在细胞中共存一段时间。子囊菌门子实层中双核的菌丝会形成一个钩状结构(crozier),以确保细胞核分裂时,两个子细胞核可以分别妥善分配到上方与下方的细胞中,维持菌丝异质双核(有来自双亲的两个细胞核)的状态。细胞中的两个核聚变之后,会立刻进行减数分裂产生子囊孢子,子囊孢子释放、散播到其他地方后可以萌发长成新的单倍体菌丝。担子菌门真菌的有性生殖过程与子囊菌门的类似,也是始于两株真菌单倍体的菌丝融合而产生双核体的菌丝,不过双核体在担子菌门更为普遍,除了子实体外,有时连营养菌丝也是双核的状态。担子菌门的子实层会形成一种称为扣子体的微观结构,与子囊菌门的钩状结构类似,可以控制细胞核分裂后两个子细胞核的分布,维持菌丝异质双核的状态。担子菌的子实体会特化出担子,担子中的两个细胞核聚变之后进行减数分裂,会产生担孢子。最为人所知的担子菌便是菇类,不过担子菌也能以其他形态存在。
接合菌门的真菌在两个体的单倍体菌丝融合后,会形成一个特化的孢子囊,孢子囊会进一步特化行成厚壁的接合孢子,对环境的抗性很大,当环境有利真菌生长时接合孢子便会萌发,进行减数分裂以产生单倍体的子细胞,并生长成菌丝。
(3)孢子的释放真菌有许多主动将无性与有性孢子释放的机制,以确保孢子可以离开子实体或菌丝而长距离传播。许多菌丝生理、机械的机制,以及孢子表面的结构都有助于孢子的散播。某些子囊菌会增加子囊中部分溶质、离子的浓度以提升膨压,造成水分流入而将子囊涨破,使子囊孢子得以释出。担子菌门的担孢子释放则与孢子表面水珠造成的表面张力有关,造成孢子弹射出的初始加速度可以高达10,000g。孢子最终弹射的距离约为0.01-0.02公分,已足以离开蕈褶,进入空气中。有些真菌孢子的释放机制较为特殊,马勃与鸟巢菌(英语:Nidulariaceae)的孢子需要雨水等外力将子实体爆破后才能释出,鬼笔科的真菌甚至具有鲜艳的颜色与特殊的气味,以吸引昆虫协助散播孢子。
(4)其他生殖方式青霉菌与麹菌有准性生殖的现象,两个个体的菌丝可能发生融合,造成菌丝中存在来自不同个体的细胞核,这些细胞核可能进一步发生核聚变,同源染色体间产生基因重组后再分裂形成单倍体的细胞,其过程与有性生殖类似,但没有减数分裂的发生,其发生频率与重要性也仍不清楚。除了在同种真菌发生外,准性生殖还可能在不同种真菌间发生,并可能是真菌演化的机制之一。
【更多精彩文章,请关注微信公众号“世界民族与文明历史”】
,




