The Scientific Research Team of HAU Has Drawn a High-Quality Fine Physical Map of the Most Complex Rye Genome.
3月18日,河南农业大学联合高校科研课题组,在国际遗传学主流期刊Nature Genetics上发布了题为“A high-quality genome assembly highlights rye genomic characteristics and agronomically important genes”的研究论文,该成果解决了长期以来悬而未决的黑麦基因组精细物理图谱组装难题,完成了中国栽培种“威宁黑麦”的基因组组装和解析,填补了作物基因组学研究重要环节。
3月18日,河南农业大学农学院植物基因组学和分子育种中心王道文研究员课题组和杨建平研究员课题组,联合河南农业大学农学院杨青华教授课题组,北京大学邓兴旺院士团队何航副教授课题组,四川农业大学任天恒副教授课题组和百迈客生物科技公司在国际顶级期刊Nature Genetics上发表了题为“A high-quality genome assembly highlights rye genomic characteristics and agronomically important genes”的研究论文,组装出目前麦类基因组组装质量最高的黑麦基因组精细物理图谱,并系统解析了黑麦的演化历史、淀粉合成、储存蛋白、抽穗期及驯化等相关基因的详细机制。同期杂志也背靠背发表了由德国莱布尼茨植物遗传与作物研究所(Leibniz Institute of Plant Genetics and Crop Plant Research,IPK)Nils Stein教授课题组牵头的黑麦基因组国际合作项目的研究成果“Chromosome-scale genome assembly provides insights into rye biology, evolution, and agronomic potential”,发布了欧洲栽培黑麦“Lo7”自交系的基因组组装结果,并解析了黑麦的演化、抗病、抗冷及自交不亲和等相关基因分子机制,建立了黑麦-小麦易位系的高通量鉴定方法,并以此为基础,深度解析了黑麦远缘易位染色体片段对小麦增量的重要作用。这些研究成果,对黑麦和小麦遗传改良具有重要的参考价值,同时也为麦类作物育种改良的源头创新提供了重要的信息资源。
☆☆☆☆☆☆☆☆☆☆☆☆☆☆☆
黑麦(Secale cereal L.)是禾本科小麦族黑麦属的二倍体植物,是小麦、大麦和燕麦的近缘物种。黑麦英文单词rye来源于古英语ryge,是“裸麦”的意思,它的中文名字可能源于其浅蓝到棕黑籽粒的色泽。与小麦和大麦一样,中东地区是黑麦的起源和野生遗传资源分布中心。黑麦具有极好的抗寒、抗病、抗逆和耐旱能力,在贫瘠土地条件下也有很强的适应能力,可耐受其他谷物无法耐受的极端气候条件,对肥料及农药需求较少,可给自然条件恶劣地区带来可观的生态和经济效益。黑麦在欧洲至少已有7,000年栽培历史,至今依然是北欧地区主要的粮食和饲料作物之一,我国西北、东北和西南山区也有种植[1]。在全球范围内,4.12百万公顷的种植面积生产了11.2百万吨的黑麦,而欧洲占全球黑麦产量的80%。在欧洲,黑麦主要集中东北地区栽培。亚洲贡献了全球13%的黑麦产量,其中50%来自中国(FAOSTAT, 2020)。

图1. 全球黑麦产量分布图
俄罗斯有名的含酒精饮料格瓦斯和大列巴面包(俄语黑麦面包хлеб的音译名字)都是以黑麦为主要原料制作而成。写进高中生物教材的“小黑麦”,是由二粒小麦(AABB)或普通小麦(AABBDD)和黑麦(RR)远缘杂交和加倍人工合成的异源六倍体或八倍体新物种。以鲍文奎院士为代表的我国老一辈科学家也在小黑麦的培育和推广上做出了杰出的贡献。相对于普通小麦和黑麦,小黑麦在生物量和抗性上优势突出,在全球范围内广泛种植(FAOSTAT,2020)。通过远缘杂交,黑麦还能替换掉小麦的染色体片段,产生稳定遗传的小麦品系,为小麦育种导入优异的外缘基因。由黑麦和小麦远缘杂交产生的1BS/1RL易位系,把黑麦的白粉病和条锈病抗性基因导入小麦,显著地提高了小麦的抗病能力和产量,为保障我国和世界粮食安全起到了至关重要的作用。目前,我国小麦品种中大约有50%以上的冬小麦品种均携带有1BL/1RS易位染色体[2]。
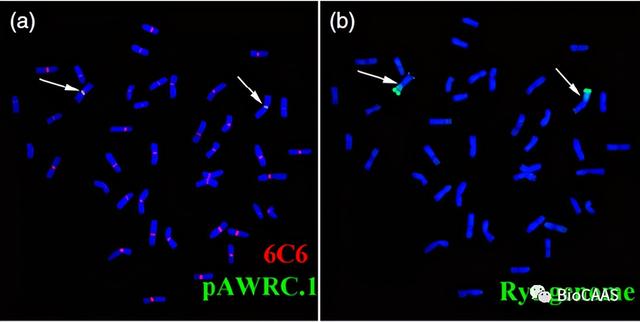
图2. 1BS/1RL易位系石4185染色体观察,箭头所示为1BS/1RL染色体(图片来自Wang et al., 2017)[3]
高质量的参考基因组,是生物学研究的基础。历时13年,耗资约30亿美元的人类基因组计划于2003年完成,被誉为是继曼哈顿原子弹计划和阿波罗登月计划之后科学史上的第三大突破。随后,水稻(2005)、玉米(2009)、大麦(2012)和小麦(2019)等重要作物的参考基因组计划也相继完成。随着技术的进步和发展,这些作物也相继进入泛基因组时代,逐渐摆脱对单一参考基因组的依赖。 相对于其他麦类二倍体基因组,黑麦基因组更加复杂(2n = 2x = 14, RR),其基因组大小估计为8 G,远大于水稻(25倍)、谷子(20倍),高粱(11倍)、玉米(4倍)、大麦(1.5倍)等其他谷物基因组,并且其基因组中有超过90%为高度重复的转座子序列。自然情况下黑麦具有自交不亲和性,属于异交作物,具有极高的杂合性,这是黑麦基因组组装的又一道很难逾越的障碍。可见,大而复杂性的黑麦基因组严重制约了麦类比较基因组学的研究、重要农艺性状遗传机制解析、作物基因学理论探讨,以及黑麦与小麦等重要作物的遗传育种改良。 为了解决上述科学问题,河南农业大学农学院联合四川农业大学、北京大学和百迈客生物科技公司等多家单位组建了协同创新研究团队,开拓性的利用中国栽培品种“威宁黑麦”强迫自交可部分结实的特性,让其强制自交18代,获得了高度纯合的自交系材料,为基因组组装扫除了部分障碍。研究团队利用高通量二代测序、PacBio单分子测序、染色质三维构象捕获 (Hi-C)、单分子光学图谱(Bionano)和高密度遗传图谱等技术,联合优化多种组装策略,成功的构建了威宁黑麦的高质量精细基因组物理图谱(图3)

图3. 威宁黑麦基因组特征分布
威宁黑麦基因组组装大小为7.84G,其中7.25G(93.67%)序列可以锚定到7条染色体上,其中5条染色体长度超过了1 Gb,最大的染色体长度(2R,1.15 Gb)为水稻基因组的3.6倍。组装得到的物理图谱与已有的欧洲黑麦(Lo7 x Lo225 RILs群体)构建的遗传图谱的相关性可达0.99。在单碱基水平通过比对发现,威宁黑麦的组装准确性可达99.99%,其基因组杂合性为0.26%。威宁黑麦基因组LAI质量参数评分为18.42,与第七版黄金标准组装的水稻参考基因组日本晴的评分近似(21.2)。其BUSCO 值为96.74%。这些结果表明,威宁黑麦基因组物理图谱得到高质量的组装完成。威宁黑麦中一共注释到45,596个高可信度基因,这些基因中有1989个抗病基因,抗病基因的数量高于大麦、水稻和其他麦类二倍体物种。 分析发现,黑麦中有6.99 Gb的转座子(transposon elements, TE),可以分为537个TE家族,占基因组比例的90.31%,显著高于其他麦类作物。长末端重复反转录转座子(LTR-RTs)对黑麦基因组扩张的贡献最大。相对于大麦,黑麦多出2.52 Gb的LTR-RTs,贡献了基因组扩张的85.42%(图4a)。黑麦基因组的扩张主要集中在几个重要的TE家族上,比如前15个TE家族约占基因组比例的56.5%,其中Daniela,Sumaya和 Sumana三个家族在黑麦中具有明显的特异性扩张(图4b)。TE演化分析发现,相对于其他二倍体麦类,黑麦中LTR插入时间具有明显的双峰分布,一次大约发生在170万年前,与大麦TE扩张时间相近;更近的一次大约发生在50万年前,与二粒小麦(AABB)形成时间和普通小麦B组祖先消亡时间近似,这是时间上的巧合还是反映了麦类演化史上一次重大的演化事件,还需进一步研究。详细研究发现,双峰分布的时间特征主要是由Gypsy超级家族的反转录转座子引起的,而Copia超级家族爆发时间距离现在更为相近(峰值为30万年)。上述分析首次系统地阐述了TE扩张与黑麦基因组演化历史的关系,如同我们仰望了很久珠穆朗玛峰,这一次我们终于攀上珠峰,亲眼目睹了其芳容。

通过与禾本科其他已测序基因组相比,在黑麦中一共鉴定到2,517个单拷贝基因。利用这一信息重新构建了禾本科系统演化史,分析发现黑麦与小麦、大麦共同祖先的分化时间大致分别在960和1,500万年前(图5a)。水稻基因组很好地保持了禾本科物种12条古染色体的基本特征,以水稻基因组为参考基因组分析发现,黑麦中有很多特有的染色体大片段重排易位事件(图5b)。但是,与水稻和大麦类似,黑麦物种的形成过程并没有经历新的全基因组复制过程。本研究得到的黑麦高质量精细基因组物理图谱,是已知组装完成最复杂的二倍体禾本科物种。该项目填补了禾本科基因组的重要空缺,为禾本科基因组演化、麦类比较基因组学的研究、重要农艺性状功能解析等提供了重要的基因组信息资源。

图5. 黑麦演化分析(a)与黑麦和水稻(b)、小麦(c)的基因组共线性
庞大的基因组往往伴随着各种基因复制现象,本研究在威宁黑麦基因组中鉴定到了23,753个分散复制基因(DDGs),6,659个近距复制基因(PDGs),7,077个串联复制基因(TDGs)和1,866个片段复制基因(图6a)。利用和黑麦演化距离最近的大麦基因组为参照,从分散复制基因中鉴定出来10,357个基因复制是由转座子引起的(TrDGs)。有意思的是,在黑麦淀粉通路合成相关基因(SBRGs)中,分别发生了5次转座复制,1次串联复制,1次分散复制和两次近距复制事件(图6d)。尤其是ScSuSy2的两个复制基因之间的表达谱发生了明显的差异,预示着它们可能产生了新的功能分化。这一发现,说明基因复制能为基因新功能分化的产生提供新的资源,从而为物种在不同条件下的适应性演化提供必要基础。这也在一定程度上揭示了黑麦突出的抗性和独特适应能力的遗传基础。

图6. 黑麦的基因复制与它们对SBRGs的多样性影响
种子储存蛋白(SSP)对麦类作物食用和加工品质具有决定性的影响。黑麦中一共有四个储存蛋白基因位点(Sec1-Sec4)。SSP位点,尤其是醇溶蛋白位点一般包含多个(最多可达几十个)高度相似的串联重复基因,导致其难以组装出完整序列,这对研究造成很大困难。本研究利用三代长片段读长的组装策略,结合三代全长转录组CCS数据辅助组装,成功实现了对这些复杂位点的组装,解析了这些基因的详细组成和结构特点(图7a)。威宁黑麦Sec-1位点长度为12 Mb,包含多个- secalins 和 -secalins串联重复基因。Sec-4位点和Sec-1位点的基因保持了很好的局部共线性,而Sec-1位点的基因和其他麦类在染色体水平保持了更好的共线性,这说明了Sec-1位点更加古老,是原始位点,而Sec-4是由Sec-1片段复制而来(图7b)。Sec-3 位点含有两个HMW-secalins 基因,和其他麦类Glu1位点保持了很好的共线性,说明高分子量麦谷蛋白在演化上具有很强的保守性。令人意外的是Sec-2位点,之前研究认为,它是由麦类作物第六同源群上的Gli2位点易位到黑麦2R染色体上,而利用高质量的物理图谱进行共线性分析却发现,黑麦的Sec-2位点除了插入的75 kb -secalins串联重复基因外,这一区域与其他麦类作物第三同源群保持了很好的共线性(图7b),这就排除了Sec-2是由Gli2位点易位而来的可能。1BS/1RL易位系提高了小麦的抗病抗逆能力和产量,但是由于1RS上Sec-1位点携带的黑麦储存蛋白的负向效应,导致小麦面粉品质下降。分析发现,Sec-1位点中缺少小麦中的低分子量谷蛋白基因,这可能是影响小麦面粉品质的主要原因之一。该研究成功解析了黑麦高度复杂的种子储存蛋白位点基因组组成,并以此为基础阐释了这些位点的演化历程,为黑麦、小麦的品质改良和产量提高打下坚实的基础。

图7. 黑麦储存蛋白位点特征(a)与共线性分析(b)
在本研究中我们也发现,“威宁黑麦”相对于“荆州黑麦”在长日照条件下抽穗期提前10-12天,与此相应的威宁黑麦中ScFT基因也会提前表达(图8a,b和c),检测ScFT蛋白发现其分子量大小(29 kDa)大于其预测大小(19 kDa),推测其可能有翻译后磷酸化修饰现象,蛋白磷酸化验证证实了这一假设(图8d和g)。该研究更进一步的对两个保守的磷酸化位点进行突变(S76D, T132D),转化烟草发现这两个位点的磷酸化消除会引起烟草植株的开花延迟,植株生物量变大(图8e,f和g)

图8. 黑麦抽穗期相关基因ScFT分析
苏联著名的遗传学家瓦维洛夫对黑麦进行了大量研究。之前的研究发现栽培黑麦是从它的野生种瓦维洛夫黑麦(S. cereale subsp. vavilovii)中驯化而来,黑麦的驯化可能来自一次必然的“意外”。其最开始可能是小麦和大麦农田中的杂草,它形态上类似于小麦、大麦栽培作物,从而避免被当做“杂草”而移除。这种行为与动物的拟态保护类似,称为“瓦维洛夫拟态(Vavilovian mimicry)”。随后,由于黑麦突出的抗寒、抗病、抗逆和耐旱能力,尤其是在贫瘠土地上出色的产出能力,让它在伴随着大麦和小麦的传播,逐步地得到驯化,从而成为一种特有的麦类作物。 利用已有的自然群体简化基因组数据和威宁黑麦高质量的参考基因组,我们分析了黑麦的全基因组驯化位点(图9a),这些区域中包含了已知的驯化或重要农艺性状的相关基因,如ScBC1, ScBtr, ScGW2,ScMOC1, ScID1 and ScWx。已知ScBC1 和 ScMOC1影响作物茎秆强度和植株形态,ScGW2和ScWx影响粒型和种子淀粉质量,ScBtr影响麦类断穗,是作物驯化的一个重要位点。ID1位点在水稻和玉米中都被证实和抽穗期相关,黑麦中其所在的6RS区间通过三种方法都能鉴定到选择信号。通过F2遗传群体,也能证实其确实是影响黑麦抽穗期差异的一个QTL位点(图9f)。进一步分析发现,黑麦中ID1位点包含了一对串联重复基因,这两个基因在编码区完全相同,而其他近缘物种只有一个基因(图9d)。这些结果解析了栽培黑麦驯化的基因组基本特征,并为黑麦独特的瓦维洛夫拟态演化假说提供了分子证据,为驯化相关基因的分子机制解析奠定了良好的基础。

图9. 黑麦栽培驯化全基因组分析
本研究综合利用多种方法和策略,克服复杂基因组组装的多重障碍,成功构建了威宁黑麦的高质量精细基因组物理图谱,并解析了其基因组的基本特征和重要农艺性状的相关分子机制。填补了作物基因组一个重要的缺失环节,为黑麦驯化、麦类基因组演化和比较基因组研究提供了重要资源。同时也将促进黑麦、小麦等相关麦类作物的遗传育种改良。 该研究由河南农业大学联合北京大学、四川农业大学、中国科学院遗传发育研究所、德国IPK莱布尼兹研究所和捷克生物技术与农业研究中心等多家单位协同完成。河南农业大学李广伟博士、王立建博士、杨建平研究员、北京大学何航副教授、河南农业大学靳怀冰博士、百迈客生物科技公司李绪明、四川农业大学任天恒副教授为本论文共同第一作者,河南农业大学王道文研究员、张坤普研究员、杨青华教授和杨建平研究员为本文章的共同通讯作者。北京大学研究生韩雪,河南农业大学研究生郑楠楠、王钊辉、张梦兰、杨书玲及师翠兰博士、熊子君博士、苟明月研究员、郑旭研究员,遗传发育所研究生赵晓歌、李峰博士、董伶俐博士、李义文博士,四川农业大学任正隆教授、颜泽洪教授、研究生宋中平和孙广华也参与了本项研究。德国IPK莱布尼兹研究所Nils Stein教授,捷克生物技术与农业研究中心Jaroslav Doležel教授,北京大学邓兴旺教授对本项目开展提供了相应的帮助。本项目得到了科技部国家重点研发计划(2016YFD0100500)、国家自然科学基金(91935304)、河南省粮食作物协同创新中心建设经费、河南省中原博新人才计划(李广伟)和全球可持续发展植物工具开发(J. Doležel)等项目的支持。参考文献:1.Schreiber, M., Himmelbach, A., Börner, A. and Mascher, M. Genetic diversity and relationship between domesticated rye and its wild relatives as revealed through genotyping-by-sequencing. Evol. Appl. 12, 66-77 (2019).2.周阳, 何中虎, 张改生, 夏兰琴, 陈新民, 高永超, 井赵斌, 于广军. 1BL/1RS易位系在我国小麦育种中的应用. 作物学报, 531-535 (2004).3.Wang J, Liu Y, Su H, Guo X and Han F. Centromere structure and function analysis in wheat-rye translocation lines. Plant J. 91, 199–207 (2017).

河南农业大学“植物基因组学和分子育种” 研究团队简介
河南农业大学“植物基因组学和分子育种” 研究团队组建于2017年4月,其主要研究方向是深入解析植物重要生命过程以及小麦等作物关键农艺性状控制的分子机理,发掘具有科学意义和育种实用价值的基因与优良变异,创制具有实用价值的新种质和新品种,为促进小麦等作物的遗传改良作出基础性、战略性、前瞻性和实用性贡献。
团队现有5位教授(王道文、杨建平、苟明月、郑旭、姬祥),其中国家杰出青年基金获得者1名。团队拥有实验室面积1440平方米,建有基因组学、分子生物学、分子育种等设施。
欢迎国内外朋友前来学术交流、合作研究。让我们携手再创辉煌!

研究团队所在的河南农业大学龙子湖校区第一实验楼

教学楼

图书馆

行政楼

龙子湖校区

教学楼
(文章转载自:BioCAAS)
统筹 EXECUTIVE
周红飞
监制 PRODUCER
陈 玺 李洪振
责任编辑 RESPONSIBLE EDITOR
邢福祥
校对 PROOFREADER
王菲凡
排版 EDITOR
王钰皓 王贺贺
来源 SOURCE
BioCAAS
,